| |
|
|
| |
Эпилептиформная активность и ЭЭГ - основные принципы
|
|
|
Эпилептиформная активность, иктальная и интериктальная, довольно разнообразна по морфологии,
топографическому распределению и динамике развития.
|
|
|
|
|
|
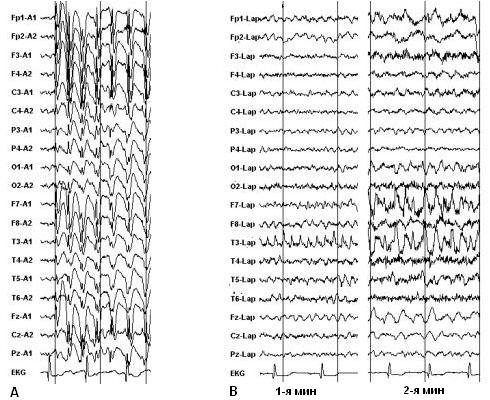
Рис. 1. А. Первично генерализованный приступ, 3-Гц комплексы спайк-волна с типичным билатерально-фронтальным максимумом амплитуды.
B. Фокальный приступ, фрагменты 1-й и 2-й минут иктальной ЭЭГ. Несмотря на постепенное вовлечение других участков мозга,
четко определяется фокус в левой височной области.
|
|
|
Независимо от того,
является эпилептиформная активность
генерализованной или фокальной, в ее
основе на клеточном уровне лежит
пароксизмальный сдвиг мембранного потенциала
(paroxysmal
depolarization
shift – PDS),
который вызывает вспышку потенциалов
действия, за которыми следует
длительная фаза гиперполяризации.
|
|
|
|
|
|
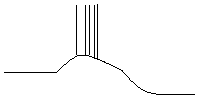
Рис.2 Пароксизмальный сдвиг
мембранного потенциала со вспышками ПД и
последующей гиперполяризацией (схема).
|
|
|
Для генерации клинически или электрографически значимой эпилептиформной активности требуется вовлечение довольно значительного количества нейронов. Предположительно выделяют два типа нейронов, играющих разную роль в этом процессе. Тип 1 – это собственно «эпилептические» нейроны, которые могут выдавать вспышки ПД самостоятельно. Тип 2 – окружающие нейроны, которые находятся под нормальным афферентным контролем, однако могут легко синхронизироваться клетками 1-го типа и вовлекаться в разряд. Вероятность вовлечения клеток 2-го типа возрастает при снижении уровня нормальной афферентации, например, в состоянии дремоты, сна.
При первично-генерализованных приступах почти мгновенно вовлекаются все области коры головного мозга. Большинство нейронов начинают работать синхронно – вспышки потенциалов действия чередуются с периодами гиперполяризации – своеобразного «электрического молчания». ЭЭГ характеризуется билатерально-синхронными комплексами спайк-волна. Начиная с 50-х гг. выдвигались различные теории, которые придавали основное значение в механизме генерализованных приступов глубинным структурам (в первую очередь, таламус), коре или обоим. Первая теория, выдвинутая в 1953, называлась «центрэнцефалической», поскольку предполагала, что эпилептиформная активность генерируется срединными глубинными структурами, достигает коры, модулируясь при этом подкорковыми структурами, в частности, таламусом. В 1968 была выдвинута «кортикоретикулярная» теория, согласно которой решающее звено принадлежит таламокортикальной «петле». Эта теория связывает первично-генерализованные приступы и механизм генерации веретен сна. На сегодняшний день в упрощенном виде модель выглядит следующим образом: первоначально возникают изменения в коре, затем вовлекаются ядра таламуса, через реципрокные таламокортикальные соединения активность усиливается, модулируется в ритм 3 Гц, трансформируясь в комплексы спайк-волна с почти мгновенной генерализацией по всей коре головного мозга
(подробнее...). Несколько по иному развиваются события при
фокальных (парциальных) приступах. Вначале формируется относительно небольшая зона
нейронов (фокус), которые переходят в пароксизмальный или «эпилептический» режим.
Запись глубинными электродами
регистрирует быструю ритмическую
активность в области фокуса.
Внутриклеточные исследования показывают,
что каждый нейрон в отдельности
работает в таком режиме относительно
недого (1-2 секунды) , ритмическая
активность представляет собой суммарный
эффект регистрации довольно большой
популяции нейронов. Постепенно на протяжении нескольких секунд, десятков секунд или минут вовлекаются все большие
участки подкорковых структур и коры, иногда – вплоть до вторичной генерализации.
При этом амплитуда колебаний нарастает,
частота уменьшается, морфологически
эпилептиформная активность представлена
ритмическими острыми волнами или
комплексами спайк-волна. Частота
колебаний продолжает снижаться, что
свидетельствует о возрастании степени
синхронизации, через какое-то время
приступ внезапно прекращается. Такую
последовательность изменения амплитуды
и частоты называют ритмической иктальной
транcформацией.
Наиболее характерным и часто встречающимся эпилептиформным паттерном являются комплексы
спайк-волна, которые могут быть локальными или генерализованными. Логотип Международной
противоэпилептической лиги представляет из себя стилизованный комплекс спайк-волна
 . В действительности
спайк-волна скальповой ЭЭГ – это
некий фантом, поскольку
при сопоставлении скальповой ЭЭГ и
интракортикальной регистрации
выясняется, что спайку соответствуют
вспышки потенциалов действия, а волне –
отсутствие всякой активности («молчание»). . В действительности
спайк-волна скальповой ЭЭГ – это
некий фантом, поскольку
при сопоставлении скальповой ЭЭГ и
интракортикальной регистрации
выясняется, что спайку соответствуют
вспышки потенциалов действия, а волне –
отсутствие всякой активности («молчание»).
|
|
|
|
|
|
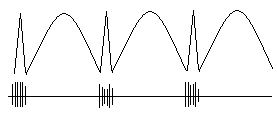
Рис. 3 Одновременная регистрация (схема) иктальной эпилептиформной активности скальповыми и кортикальными электродами.
|
|
|
Таким образом, вспышки потенциалов действия – это «позитивный» (усиленная, чрезмерная активность) феномен, которому соответствует спайк или острая волна в скальповой ЭЭГ. Последующая медленная волна – это «негативный» феномен, поскольку ей соответствует отсутствие активности корковых нейронов. Медленная волна, которую мы видим в скальповой ЭЭГ, является, таким образом, скорее феноменом регистрации, а не генерации эпилептиформной активности. Оговоримся еще раз, что это касается медленной волны, как компонента комплекса спайк-волна.
Отдельного упоминания заслуживает электродекремент – кратковременное падение амплитуды фоновой активности на скальповой ЭЭГ, которое может предшествовать приступу. Длительность, как правило, в пределах 1-2 секунд. Электродекремент может быть локальным или регистрироваться по всем отведениям. На сегодняшний день нет общепринятого объяснения этого феномена. Одно из предположений сводится к тому, что электродекремент есть следствие активации восходящей ретикулярной системы, которая запускает эпилептический приступ. Иногда электродекремент является единственным регистрируемым изменением в скальповой ЭЭГ во время приступа. Кратковременное падение амплитуды наиболее характерно для инфантильных спазмов.
|
|
|
|
|
|
|
|
|
Подготовил А.Г.Брутян
|
|
|
Список литературы
|
|
|
Avoli M, Gloor P, Kostopoulos G, Gotman J. An analysis of penicillin-induced generalized spike-and-wave discharges using simultaneous recording of cortical and thalamic single units. J Neurophysiol 1983; 50:819-37.
/Medline/
Binnie CD, Stefan H. Modern Electroencephalography: its role in epilepsy
management. Clin Neurophysiol 1999, 110: 1671-97. /Medline/
Bancaud J, Talairach J, Geier S, Bonis A, Trottier S, Manrique M. Behavioral manifestations induced by electric stimulation of the anterior cingulate gyrus in man. Rev Neurol (Paris) 1976; 132:705-24.
/Medline/
Geiger LR, Harner RN. EEG patterns at the time of focal seizure onset.Arch Neurol 1978; 35: 276-86.
Gloor P. Generalised cortico-reticular epilepsies. Some considerations on the pathophysiology of generalised bilaterally synchronous spike and wave discharge. Epilepsia 1968; 9:249-263.
Halasz P. Generalized epilepsy with spike-wave paroxysms as an epileptic disorder of the function of sleep promotion. Acta Physiol Acad Sci Hung. 1981; 57(1):51-86.
/Medline/
Marcus EM, Watson CW, Simon SA. An experimental model of some varieties of petit mal epilepsy: electrical-behavioral correlations of acute bilateral epileptogenic foci in cerebral cortex. Epilepsia 1968; 9:233-248.
Pollen DA, Perot P, Reid KH. Experimental bilateral wave and spike from thalamic stimulation in relation to level of arousal. Electroenceph clin Neurophysiol 1963; 15:1017-1028.
Stefan H, Snead OC. In: Engel J, Pedley TA, editors. Epilepsy: a comprehensive textbook, Philadelphia, PA: Lippincott-Raven, 1997: 79-590.
Williams D. A study of thalamic and cortical rhythms in petit mal. Brain 1953; 76: 50-69.
Wyler AR, Fetz EE, Ward AA. Antidromic and orthodromic activation of epileptic neurones. Experimental Neurol 1974; 43: 59-74.
/Medline/
Wyler AR, Fetz EE, Ward AA. Spontaneous fring patterns of epileptic neurones in the monkey motor cortex. Experimental Neurol 1973; 40:567-585.
/Medline/
|
|
|
|
|
|
|
|
|
